Der schnellste Pfad der Evolution
Forschungsbericht (importiert) 2007 - Max Planck Institut für Evolutionsbiologie
Motivation: Kugelbahnen und Physik
Die Mathematisierung der Biologie ist im Vergleich zur Physik noch recht jung. Nur wenige allgemeine Grundsätze, wie z.B. das Hardy-Weinberg-Gesetz für sexuelle Reproduktion, sind bekannt. Solche Gesetzmäßigkeiten kann man an vereinfachten, abstrakten Modellen finden. In der Physik haben diese Modelle eine lange Tradition.
Ein klassisches Problem der theoretischen Physik ist das so genannte Brachistochronenproblem [1]: Man kann hierbei z.B. an eine Kugelbahn zwischen zwei Punkten A und B denken. Welche Form muss nun eine solche Kurve haben, damit die Kugel möglichst schnell Punkt B erreicht? Ist die Kurve am Anfang zu steil, dann wird die Kugel zwar stark nach unten beschleunigt, legt aber nur eine sehr geringe horizontale Entfernung zurück. Ist die Kurve zu flach, beginnt die Bewegung nur äußerst langsam.
Johann Bernoulli forderte die Wissenschaftsgemeinde 1696 in einer Veröffentlichung in Acta Eruditorium auf, dieses Problem zu lösen. Neben Johann Bernoulli fanden sein Bruder Jacob, Guillaume de l’Hopital, Gottfried Leibniz und Isaac Newton eine Lösung: Die Kurve, auf der die Kugel am schnellsten herunterrollt, ist eine Zykloide – die Kurve, die ein Punkt auf einem rollenden Rad beschreibt (Abb. 1, oben).
 wird durch die angegebene Gleichung mit dem Parameter t beschrieben. Unten: Wenn man das Problem umdreht, kommt man zu einem verwandten Problem der Evolutionsbiologie: Wie muss man Fitnesswerte ri anordnen, um durch Selektion möglichst schnell die Fitness auf einen bestimmten Wert R zu erhöhen? Für kleine Mutationsraten muss die Kurve exponentiell ansteigen, wie aus der Gleichung ersichtlich.")
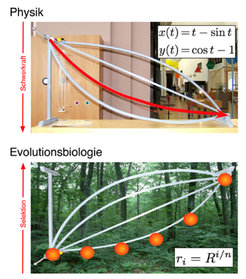
Kugelbahnen in der Evolutionsbiologie
Gibt es eine ähnliche Fragestellung in der Evolutionsbiologie?
Eine Grundfrage in diesem Forschungsgebiet ist, auf welche Weise sich vorteilhafte Eigenschaften durch die natürliche Selektion in Populationen durchsetzen. Die theoretische Biologie entwickelt hierzu mathematische Modelle, mit denen die Eigenschaften solcher Prozesse untersucht werden können. Wichtige Merkmale solcher Prozesse sind zum Beispiel die Wahrscheinlichkeit, dass sich vorteilhafte Eigenschaften durchsetzen oder die Dauer dieses Prozesses.
Wenn man nun an die Kugelbahnen zurückdenkt, dann tritt bei Evolutionsprozessen die natürliche Selektion an die Stelle der Gravitation. Statt durch Höhenunterschiede wie in der Physik wird der Prozess durch Unterschiede in der Fitness getrieben, wobei in der Physik die Höhe (bzw. die potenzielle Energie) abnimmt und in der Biologie die Fitness zunimmt. Dies führt zu einem biologischen Optimierungsproblem, das mit dem Brachistochronenproblem verwandt ist [2]. Wie muss man die Fitnesswerte auf einer Kurve über mehrere Zwischenzustände wählen, damit eine Population möglichst schnell von einer Ursprungsfitness zu einer Zielfitness evolviert (Abb. 1, unten)?
An dieser Stelle zeigt sich, dass das biologische Problem nicht ganz so elegant zu lösen ist wie das physikalische. Während die Lösung in der Physik von keinen Parametern mehr abhängt, beeinflusst in der Biologie die Mutationsrate das Ergebnis. Der Einfachheit halber kann man annehmen, dass es immer nur eine einzige vorteilhafte Mutation gibt, d.h. die vorteilhaften Eigenschaften müssen nach und nach in einer bestimmten Reihenfolge akkumuliert werden. Weiter muss man definieren, wie sich vorteilhafte Eigenschaften in einer Population durchsetzen. Ein Standardansatz der Populationsgenetik dazu ist der Moranprozess [3]: In jedem Zeitschritt wird ein Individuum zufällig, aber proportional zur Fitness ausgewählt. Dieser Vorgang entspricht einem Roulettespiel, bei dem jedes Feld ein Individuum repräsentiert. Die Größe der Felder ist durch die Fitness gegeben. Das so ausgewählte Individuum produziert einen identischen Nachkommen. Anschließend wird ein zufälliges Individuum unabhängig von der Fitness aus der Population entfernt, um die Populationsgröße konstant zu halten. Auf diese Weise produzieren die Individuen mit höherer Fitness mehr Nachkommen und setzen sich mit einer höheren Wahrscheinlichkeit durch. Da es sich jedoch um einen Zufallsprozess handelt, kann es auch sein, dass ein bestimmter Typ von Individuen verloren geht, obwohl er vorteilhaft ist. Dieser Prozess führt über kurz oder lang immer zu einer Population, in der alle Individuen gleich sind. Man kann zusätzlich die Möglichkeit berücksichtigen, dass bei der Reproduktion Fehler oder Mutationen auftreten, die den Fitnesswert verändern. So kann man untersuchen, ob sich diese neuen Individuen in der Population durchsetzen.
Wie muss man nun die Fitnesswerte wählen, um eine möglichst schnelle Evolution zu erreichen? Im allgemeinen Fall kann selbst dieses einfache Problem nicht mit Papier und Bleistift gelöst werden – hier können Computersimulationen hilfreich sein. In Grenzfällen, die hingegen schwieriger mit Simulationen untersucht werden können, sind jedoch analytische Näherungen möglich.
Biomathematik mit Bleistift und Papier
Falls Fehler oder Mutationen bei der Reproduktion sehr unwahrscheinlich sind, dauert es sehr lange, bis alle Mutationen aufgetreten sind. In diesem Grenzfall setzt sich eine Mutante immer durch oder geht verloren, bevor die nächste Mutation stattfindet. Die Population besteht also fast immer aus gleichen Individuen, und ein Beobachter würde die meiste Zeit auf Mutationen warten. Nur selten tauchen Mutationen auf, die entweder wieder aussterben oder sich in der Population durchsetzen.
In diesem Fall zeigt eine einfache Rechnung, dass die Fitnesswerte im Idealfall exponentiell ansteigen (Abb. 1, unten). Weiter kann man mathematisch beweisen, dass diese Lösung eindeutig ist – es kann also keine alternative Lösung des Problems geben, die genauso gut ist.
Dazu ein Beispiel: Bei R=16 und drei Zwischenzuständen heißt das also, dass die Fitnesswerte 1,2,4,8,16 sein sollten. Im Fall von sehr vielen Zwischenzuständen liegen diese auf der einzigen exponentiell ansteigenden Kurve, die Anfangs- und Endzustand verbindet.
Dieses einfache Ergebnis gilt jedoch nur in dem Grenzfall, in dem die Mutationsrate sehr klein ist. Genauer gesagt, muss sie viel kleiner sein als die inverse Populationsgröße zum Quadrat. Wenn also z.B. die Population aus 100 Individuen besteht, muss die Mutationsrate kleiner als 0.0001 sein. Diese Näherung ist also nur dann gültig, wenn die Population nicht allzu groß ist.
Man kann jedoch eine andere Näherung durchführen, die genau dann gültig wird, wenn die Population groß ist und die Mutationsrate hoch. In diesem Fall läuft der gesamte Prozess sehr schnell ab, wenn man ihn mit niedrigen Mutationsraten vergleicht. Für hohe Mutationsraten besteht die Population sehr schnell aus allen möglichen verschieden Typen von Individuen. Insbesondere sind Individuen vom Typ mit der maximalen Fitness vorhanden. Ein Beobachter würde in diesem Fall nur darauf warten müssen, dass sich diese Individuen durchsetzen. Anstatt wie für niedrige Mutationsraten die Wahrscheinlichkeit, dass sich eine Mutation durchsetzt zu optimieren, muss man nun an der Zeit ansetzen, die dieses Durchsetzen dauert. Dieser Prozess ist am effektivsten, wenn der Fitnessvorteil dieser Individuen maximal wird. Und er ist genau dann am schnellsten, wenn alle Individuen die Anfangsfitness haben und nur der finale Typ eine höhere Fitness. Die schnellste Trajektorie, also der schnellste Pfad, ist in diesem Falle also flach mit einem Maximum ganz am Ende (Abb. 2). Diese Näherung ist dann gültig, wenn das Produkt aus Populationsgröße und Mutationsrate groß ist. Im allgemeinen Fall erwartet man, dass es einen kontinuierlichen Übergang von der exponentiellen Trajektorie zur flachen Trajektorie gibt. Computersimulationen bestätigen dies.
. Sind dagegen die Mutationsraten hoch und die Populationen groß, dann ist der schnellste Pfad flach mit einem großen Fitnesssprung zum finalen Zustand (rechts). Dies sagt jedoch nichts über die absoluten Geschwindigkeiten des Prozesses aus: In jedem Fall wird die Evolution schneller voranschreiten, wenn die Mutationsraten hoch sind.")
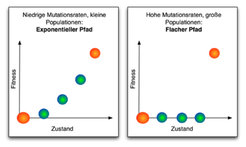
Für hohe Mutationsraten ist die absolute Zeit des Prozesses immer kürzer als für niedrige Mutationsraten. Etwas Ähnliches ist ganz alltäglich: Die schnellste Strecke mit dem Fahrrad kann sehr verschieden sein von der schnellsten Strecke mit dem Auto, obwohl die gesamte Fahrzeit mit dem Auto immer kürzer ist. Auf ähnliche Weise, wie in diesem Beispiel das Verkehrsmittel vor der Optimierung festgelegt wird, bestimmen in dem Modell die Mutationsraten, welche Form der optimale Pfad hat.
Gibt es so etwas auch in der Natur?
Ist dieses Problem nun relevant für die experimentelle Biologie? Dazu zuallererst eine Gegenfrage: Für wen ist das Brachistochronenproblem der Physik interessant? Obwohl sich vielleicht Skifahrer dafür erwärmen könnten, ist es doch weit weg von echten Anwendungen. Trotzdem war die Lösung des physikalischen Problems ein wichtiger Schritt in der Entwicklung der theoretischen Mechanik und führte zum Beispiel zur Entwicklung der Variationsrechnung, einer neuen mathematischen Methode in der Physik.
Normalerweise wird in der Evolution jedoch folgendes beobachtet: Zuerst steigt die Fitness sehr schnell an, später gibt es nur noch sehr wenige Verbesserungsmöglichkeiten und die Fitness steigt nur noch langsam an. Trotzdem ist die Theorie mit der Realität kompatibel: auch eine Kugel wird anfangs in die Richtung rollen, in der das Gefälle am größten ist – diese „steilster Abstieg“-Methode wird auch in der Optimierungstheorie verwendet. Die Berechnung des schnellsten Pfades sagt aber nichts über den realisierten Weg aus: Auch wenn es einen schnellsten Pfad gibt, heißt das noch lange nicht, dass die Evolution diesen Weg auch beschreitet.
Solche Ketten von Mutationen gibt es in unterschiedlichsten Systemen. Krebs kann durch die Inaktivierung von Tumor-Supressor-Genen initiiert werden, dabei ist die erste Mutation neutral, da wir zwei Kopien des Gens besitzen. Erst die zweite Mutation führt zu einer erhöhten Fitness der Krebszellen (die für den Menschen natürlich von Nachteil ist) und zu dem Ausbruch der Krankheit [4,5]. Pathogene müssen oftmals mehrere Mutationen ansammeln, um den Wirt verlassen zu können [6]. In der experimentellen Evolution kann bisweilen die Fitness von Zwischenzuständen manipuliert werden. Auf diese Weise kann man die hier dargestellten theoretischen Ergebnisse auch experimentell überprüfen. Vergleicht man zwei Systeme, dann wird das System schneller den Endzustand erreichen, das dichter an der optimalen Trajektorie liegt.
In der Evolutionsbiologie geht man oft davon aus, dass die Fitness bereits maximal ist. Wenn die Evolution aber nur über einen geringen Zeitraum fortschreitet (so wie bei der experimentellen Evolution oder bei der Evolution von Krebs), dann wird der Prozess der Evolution selber wichtig und nicht nur dessen Endergebnis. Für Fragestellungen wie die hier diskutierten sind theoretische und mathematische Biologie ausgezeichnete Hilfsmittel.