Diversität und nachhaltige Nutzung amazonischer Überschwemmungswälder
Forschungsbericht (importiert) 2006 - Max Planck Institut für Evolutionsbiologie
Einleitung
Der nachhaltige Schutz tropischer Regenwälder ist eine wesentliche Voraussetzung für den Erhalt der genetischen Vielfalt der Erde und der Stabilität des globalen Klimas. Nach heutigem Wissensstand beherbergen tropische Regenwälder mehr als 50.000 Baumarten und haben einen Anteil von 32 bis 36 % in der globalen Bilanz der Pflanzen-Nettoprimärproduktion. Mit einer Fläche von 6 Mio. km2 stellt das Amazonasbecken den größten zusammenhängenden tropischen Regenwald dar. Amazonische Tiefland-Regenwälder bestehen aus einer Vielzahl unterschiedlicher Ökosysteme, die sich u.a. durch eigene Pflanzengesellschaften voneinander abgrenzen. Periodische Überschwemmungsgebiete entlang der großen Flussläufe Amazoniens bedecken hierbei eine Fläche von ca. 300.000 km2.
Aufgrund der leichten Zugänglichkeit und des hohen Nährstoffangebots in den Schwemmlandböden sind die Überschwemmungsgebiete von Weißwasserflüssen (várzea) die am dichtesten besiedelten Bereiche Amazoniens mit hohem Potenzial für Land-, Vieh- und Holzwirtschaft sowie für Fischerei. Die intensive landwirtschaftliche Nutzung und die größtenteils unkontrollierte Exploitation ihrer Nutzhölzer führte jedoch zur großflächigen Reduzierung von Primärwäldern und zu lokalen Artenverlusten insbesondere im dichter besiedelten östlichen Amazonien.
Diversität amazonischer Überschwemmungswälder
Nahezu alle Pflanzenfamilien der tropischen Flora Lateinamerikas sind auch in amazonischen Überschwemmungswäldern vertreten. Mit mehr als 900 bisher bekannten Baumarten ist die várzea der artenreichste Überschwemmungswald der Erde und somit ca. 20-mal artenreicher als die Überschwemmungswälder Mitteleuropas sowie 5- bis 10-mal artenreicher als subtropische und tropische Überschwemmungswälder Asiens und Afrikas. Hauptgründe für den hohen Artenreichtum der várzea sind hohe Habitatdiversität, seit dem frühen Tertiär relativ stabile tropisch-subtropische Klima-, Überflutungs- und Wachstumsbedingungen, welche evolutive Anpassungen der Arten an die alljährliche Überschwemmung und somit hohe ökologische Einnischung fördern, sowie der stetige Artenaustausch mit den umgebenden artenreichen Wäldern auf nicht überschwemmten Standorten (terra firme).
Várzea-Wälder bestehen aus unterschiedlichen Pflanzengemeinschaften und Waldtypen, die sich in natürlicher Sukzession entlang des Flutgradienten entwickeln. Dabei ist der „monomodale Flutpuls“ mit einer vorhersehbaren Hochwasser- und Niedrigwasserperiode im Jahresverlauf entscheidend für die Artenverbreitung und die Wachstumsstrategie von Bäumen. Nach erster Anlagerung neuen Sediments durch die Flüsse entstehen zunächst artenarme Pionierwald-Gesellschaften aus hochspezialisierten Arten, tolerant gegenüber extremer Sonneneinstrahlung und gegenüber Überschwemmungen von bis zu 7,5 Metern Höhe und einer Dauer von bis zu 230 Tagen im Jahr. Als „schattenspendende Sedimentfallen“ bewirken diese Arten ein „Höhenwachstum“ des Standorts und fördern die Etablierung von Sekundärgesellschaften, welche sich aus weniger starklicht- und überflutungstoleranten Arten zusammensetzen (várzea baixa-Wälder). Am Ende der Sukzession stehen artenreiche Schlusswald-Gesellschaften (várzea alta-Wälder), die eine zweimonatige Überflutung ihres Wurzelbereichs tolerieren, aber nicht über die spezifischen Anpassungen verfügen wie die Arten der várzea baixa-Wälder. Höchster beschriebener Baumartenreichtum (Bäume über 10 cm Brusthöhendurchmesser BHD) sind 84 Arten pro Hektar (ha-1) in Ostamazonien, 142 Arten ha-1 in Zentralamazonien, und 157 Arten ha-1 in Westamazonien. Damit entspricht der Artenreichtum der várzea etwa pro Flächeneinheit zu 60-70% dem der umgebenden terra firme (Abb. 1).
, high várzea (HV) und nicht überschwemmter Wälder (terra firme;TF) in Ost-, Zentral-, und Westamazonien im Vergleich.
Low várzea: Überschwemmungshöhe > 3 m, Überschwemmungsdauer > 50 Tage im Jahr; high várzea: Überschwemmungshöhe < 3 m, Überschwemmungsdauer < 50 Tage im Jahr. KmW: km westlich.")
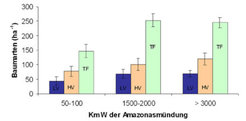
Etwa 30% aller Baumarten der várzea alta-Wälder kommen auch in der umgebenden terra firme vor. Die Anzahl gemeinsamer Arten sinkt rapide mit zunehmender Entfernung zwischen den Bestandesaufnahmen und beträgt bei 1000 km entfernten Beständen nur ca. 10%. Low várzea-Bestände beinhalten dagegen nur wenige Arten der umgebenden terra firme, können aber über tausende von Kilometern hohe floristische Similarität aufweisen. Der Grund hierfür liegt in der evolutiven Ausbildung spezifischer Anpassungen vieler low várzea-Arten an die hohen Überschwemmungen. Viele Arten verbreiten ihre Diasporen über das Wasser (Hydrochorie) und über samen- und fruchtfressende Fische (Ichthyochorie), sodass das Flusssystem als wirkungsvoller Verbreitungsvektor genutzt wird.
Zusammenhänge zwischen Flutpuls, Baumwachstum und Oberflächenwassertemperaturen des tropischen Pazifiks und tropischen Nordatlantiks
Der Wachstumsrhythmus der Bäume im Überschwemmungsgebiet wird vom Flutpuls gesteuert. Mit Beginn der aquatischen Phase wird das Holzwachstum reduziert und es kommt zu einer kambialen Ruhe und somit zur Bildung eines Jahresringes im Holz der Baumarten. Das Holzwachstum der Bäume in den Überschwemmungswäldern ist maßgeblich auf die terrestrische Phase beschränkt und die Jahresringbreiten im Holz korrelieren somit mit der Dauer der terrestrischen Phase des entsprechenden Jahres (Vegetationsperiode).
Die Niederschläge und der daraus resultierende Flutpuls im zentralen Amazonasbecken sind maßgeblich von den Oberflächenwassertemperaturen (OWT) des tropischen Nordatlantiks und tropischen Pazifiks beeinflusst. In El-Niño-Jahren fallen im Amazonasbecken insgesamt weniger Niederschläge im Vergleich zu anderen Jahren. Daraus resultiert ein abgeschwächter Flutpuls mit einer verkürzten aquatischen Phase und einer verlängerten terrestrischen Phase. Jahresringbreiten in El-Niño-Jahren sind somit signifikant breiter als in anderen Jahren. Die Zusammenhänge zwischen El Niño, Flutpuls und Baumwachstum ermöglichen die Rekonstruktion des Einflusses von El Niño auf die Überschwemmungsgebiete anhand über 200-jähriger Jahresringchronologien.
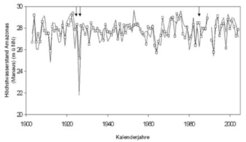
Anhand der gewonnenen Erkenntnisse über die Zusammenhänge zwischen Baumwachstum und Überschwemmung und deren Assoziierung mit ozeanographischen Daten wurde ein Modell entwickelt, um Höchstwasserstände in Zentralamazonien vorhersagen zu können. Bereits im Februar kann der im Juni auftretende Höchstwasserstand anhand des mittleren Wasserstandes des Rio Negro im Februar und des Southern Oscillation Index – ein meteorologischer Index, der die Luftdruckunterschiede zwischen Darwin (Australien) und Tahiti (tropischer Pazifik) berücksichtigt – akkurat vorhergesagt werden (Abb. 2). Niedrigwasserstände des Amazonas treten gewöhnlich in der 2. Oktoberhälfte/1. Novemberhälfte eines Jahres auf und sind vor allem mit Oberflächenwassertemperaturen (OWT) des tropischen Atlantiks der Nordhemisphäre assoziiert. Erhöhte OWT im tropischen Nordatlantik sind auch die Ursache zur Bildung von Zyklonen und Hurrikanen, welche vermehrt ab einer Oberflächenwassertemperatur von über 26 °C entstehen. Der ACE-Index (Accumulated Cyclone Energy Index), der sich aus der Häufigkeit, Intensität und Dauer der Zyklone und Hurrikane eines Jahres errechnet, zeigt einen signifikanten Zusammenhang mit den Niedrigwasserständen in Zentralamazonien. In Jahren mit hohen ACE-Indizes werden große Mengen an Feuchtigkeit in die Nordhemisphäre (Karibik, Golf von Mexiko) transportiert und es gelangt weniger Feuchtigkeit in das Amazonasbecken. Aufgrund der prognostizierten Erwärmung des tropischen Pazifiks und Nordatlantiks können zukünftig vermehrt extreme Niedrigwasserstände und Trockenheiten in Amazonien erwartet werden.
 und vorhergesagten Höchstwasserständen (Linie mit Punkten) des Amazonas bei Manaus für den Zeitraum 1903-2005. Nur in 3% der Fälle (angezeigt durch Pfeile) sind die Abweichungen zwischen vorhergesagtem und beobachtetem Wert größer als 150 cm.")
Kohlenstoffspeicherung und -aufnahme in der oberirdischen Biomasse der zentralamazonischen Überschwemmungswälder
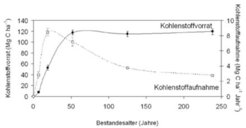
Tropische Regenwälder sind ein dynamisches Kohlenstoffreservoir, jedoch auch Quelle von Kohlenstoffemissionen im Zuge fortschreitender Rodung, insbesondere zur Flächengewinnung für Landwirtschaft und Viehzucht. In den zentralamazonischen Überschwemmungswäldern wurden die Kohlenstoffvorräte und Kohlenstoffaufnahme in der oberirdischen Holzbiomasse von Sukzessionsstadien mit Bestandesaltern zwischen 7 und 240 Jahren berechnet; dies anhand von Waldinventuren und Wachstumsmodellierungen basierend auf Jahresringanalysen. Entlang des Sukzessionsgradienten kann eine Zunahme der Kohlenstoffvorräte auf 115-120 Tonnen Kohlenstoff pro Hektar nur bis zu einem Bestandesalter von 50 Jahren beobachtet werden (Abb. 3). Kohlenstoffvorräte in der oberirdischen Holzbiomasse der älteren Waldbestände nehmen nicht zu und befinden sich im Gleichgewicht. Kohlenstoff, der von der Holzbiomasse im Zuge des Dicken- und Höhenwachstums der Bäume aufgenommen wird, geht an anderer Stelle durch Mortalität wieder verloren. Die Holzbiomasse der älteren Waldbestände der zentralamazonischen Überschwemmungswälder fungiert somit nicht als Kohlenstoffsenke.
. Nur Wälder mit Bestandesaltern bis zu 50 Jahren nehmen tatsächlich Kohlenstoff in die oberirdische Holzbiomasse auf, angezeigt durch den ansteigenden Kohlenstoffvorrat. Bei den älteren Wäldern nimmt trotz der Kohlenstoffaufnahme der Kohlenstoffvorrat nicht zu.
Mg C ha-1: Megagramm (Tonne) Kohlenstoff pro Hektar")
Nachhaltige Holznutzung in zentralamazonischen Überschwemmungswäldern
Die Nutzung der Tropenwälder nach Prinzipien der Nachhaltigkeit ist ein vielversprechender Weg, um die vielfältigen Wirkungen und Funktionen dieser Wälder zu sichern und gleichzeitig die Versorgung der lokalen Bevölkerung und Industrie mit Holzressourcen zu garantieren. Durchmesserwachstumskurven basierend auf Jahresringanalysen zeigen, dass Wirtschaftsbaumarten in der várzea zwischen 13 und 140 Jahre benötigen, um den gesetzlich vorgeschriebenen Mindesthaubarkeitsdurchmesser von 45 cm zu passieren (Abb. 4). Aus den Jahresringanalysen entwickelte Wachstumsmodelle belegen, dass Nutzungszyklen zwischen verschiedenen Baumarten und Waldökosystemen variieren. In der nährstoffreichen várzea betragen die Nutzungszyklen zwischen 3 und 31 Jahren und im nährstoffarmen igapó zwischen 37 und 53 Jahren. Diese Ergebnisse machen deutlich, dass Bewirtschaftungskonzepte in Amazonien und in den Tropen allgemein ökologisch nicht nachhaltig sind, da sie für alle bewirtschafteten Baumarten denselben Nutzungszyklus anwenden. Eine ökologisch nachhaltige Waldbewirtschaftung muss baumarten- und standortspezifisch erfolgen.
 von 12 Weich- und Harthölzern der zentralamazonischen Überschwemmungswälder der várzea unter Berücksichtigung des gesetzlich festgelegten Mindesthaubarkeitsdurchmessers von 45 cm.")
