Genetisch diverse Populationen trotzen dem Klimawandel – experimentelle Erkenntnisse aus Seegraswiesen
Forschungsbericht (importiert) 2004 - Max Planck Institut für Evolutionsbiologie
Die überwiegend positiven Auswirkungen von lokaler Artenvielfalt sind mittlerweile in zahlreichen Experimenten gut belegt [1]. Lebensgemeinschaften mit vielen Arten sind stabiler, weisen höhere Ökosystemleistungen wie Produktivität oder Nährstoffaufnahme auf und erholen sich nach Störungen schneller als artenarme Systeme. Allerdings sind viele Pflanzenbestände von Natur aus Monokulturen. Dies gilt zum Beispiel für Schilfbestände, Salzmarschen und Wasserpflanzenbestände in Seen und im Meer. Solche aquatischen Makrophyten gehören gleichzeitig zu den produktivsten Pflanzenbeständen überhaupt. Forschungen am Max-Planck-Institut für Limnologie, Plön, und dem Leibniz-Institut für Meereswissenschaften in Kiel hatten zum Ziel, diesen scheinbaren Widerspruch an Seegraswiesen der westlichen Ostsee aufzuklären [2]. Solche Wiesen bestehen ebenfalls meist nur aus einer so genannten Gründerart, von der, ganz ähnlich den Wäldern an Land, ein ganzes Ökosystem abhängt. In Ost- und Nordsee ist dies das Große Seegras (Zostera marina), eine marine Blütenpflanze, welche in Küstenzonen der gemäßigten Breiten die einzige Art ist, die unterseeische Wiesen ausbildet. Auf der Ebene von Arten besteht also keinerlei Redundanz im Falle eines Verlustes von Z. marina-Populationen. Ganz im Gegensatz zur einartlichen Pflanzengemeinschaft ist die in den Wiesen lebende Fauna sehr vielgestaltig und besteht aus Kleinkrebsen, Jungfischen, Muscheln und Schnecken (Abb. 1).
 in der süd-westlichen Ostsee (A) sowie typische Vertreter wirbelloser Tiere, die zwischen den Blättern leben: (B) Strandkrabbe Carcinus maenas (C) Schwebgarnelen Mysis spp. im Hintergrund, Meeresassel Idotea balthica frisst Aufwuchs der Seegrasblätter (Pfeil).")

Versteckte Vielfalt auf genetischer Ebene
Doch ist ein Seegrasbestand tatsächlich so uniform, wenn die Erbinformationen mit berücksichtigt werden? Molekulargenetische Marker ermöglichen es, durch einen ‚genetischen Fingerabdruck’ die Vielgestaltigkeit von Individuen und Populationen auf DNA-Ebene zu messen. Durch das Vergrößerungsglas molekulargenetischer Marker ändert sich das Bild scheinbarer Uniformität drastisch. Dann sind scheinbar monotone Seegrasbestände voller komplexer Diversität. Es lassen sich Bereiche mit zahlreichen einzigartigen Genotypen (Klonen) von solchen unterscheiden, bei denen sich bestimmte, sehr erfolgreiche Genotypen durch natürlicherweise ablaufende Klonierung über Rhizome (Erdsprosse) in der Fläche ausgedehnt haben (farbige Bereiche in Abb. 2).
Das ursprüngliche Ziel eines Experimentes war es, im Freiland erstmalig die Auswirkungen genotypischer Diversität auf verschiedene Populationsparameter wie Dichte und Biomasse zu ermitteln. Dazu stellten Thorsten Reusch und seine Mitarbeiter kontrollierte Mischungen aus einem (Monokulturen), drei oder sechs Genotypen zusammen und pflanzten sie im flachen Wasser einer geschützten Meeresbucht bei Maasholm (süd-westliche Ostsee) aus. Basis für die Experimente war eine große Klonkarte mit 30 x 30 (=900) Probennahmepunkten in einem Raster direkt neben den geplanten Experimentalflächen (Abb. 2). Die verbindenden Rhizome zwischen Blattsprossen verrotten nach wenigen Monaten, Seegrasklone werden jedoch über 100 Jahre alt. Deswegen erfolgte die Identifizierung von Genotypen mit Hilfe von Mikrosatelliten-Markern, die über die Polymerase-Kettenreaktion variable Bereiche im Erbgut sichtbar machen können.
 in Maasholm, süd-westliche Ostsee. Blattproben zur DNA-Analyse wurden im Abstand von 33,3 cm genommen. Genotypen oder Klone wurden mithilfe des Polymorphismus von neun Mikrosatelliten-Markern bestimmt. Gleiche Zahlen geben gleiche Multilokus-Genotypen an. Die acht größten Genotypen inklusive der im Experiment verwandten Klone (A-F) sind farbig eingezeichnet.")

Experimentelle Effekte genotypischer Diversität im Hitzejahr 2003
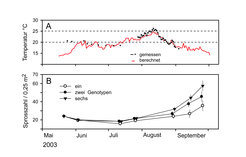
Das Experiment fiel jedoch in den extrem warmen Sommer 2003, der als Modelljahr für die vorhergesagte Zunahme von klimatischen Extremereignissen im Rahmen von ‚global change’ dienen kann [3]. Im Flachwasser der Ostsee wurden Wassertemperaturen von über 25°C gemessen, die für viele winterkalte und kaltgemäßigte Organismen an der Grenze der Temperaturtoleranz sind, so auch für Z. marina. Somit ergab sich die einzigartige, ungeplante Möglichkeit, die Antwort von Populationen auf die vorhergesagten klimatischen Extremereignisse im Experiment zu verfolgen. Etwa die Hälfte der Seegraspflanzen starben ab. Erst im Spätsommer erholten sich die Bestände, wobei sich die genotypisch diversen Experimentalflächen signifikant schneller erholten [2], Abbildung 3. Auf die schnellere Erholung der Seegrasflächen reagierte auch die assoziierte Fauna von Krebsen, Schnecken und juvenilen Muscheln mit höheren Dichten, jedoch nicht mit einer größeren Diversität. Auch sie zeigte deutlich höhere Dichten auf den genetisch vielfältigen Experimentalflächen mit sechs Genotypen.
 sowie die Entwicklung der mittleren Sprosszahlen (± ein Standardfehler) in Abhängigkeit von der genotypischen Diversität (B). Die Wassertemperaturen wurden teilweise direkt am Experimentalstandort gemessen oder aber aus Temperaturdaten in ca. 50 km Entfernung berechnet.")
Echte und ‚unechte’ Effekte genetischer Vielfalt
Die positive Wirkung von genotypischer Diversität kann mit zwei Hauptmechanismen zusammenhängen: Selektionseffekten und Komplementarität. Selektionseffekte kommen zustande, wenn in diverseren Mischungen aus statistischen Gründen solche Genotypen die Populationsleistung dominieren, die unter genau den lokalen Bedingungen am besten wachsen. Da dies eine unausweichliche Konsequenz verschieden großer Stichproben von Genotypen ist, kann man hier schwerlich von einem Effekt von Vielfalt per se sprechen. Komplementarität dagegen wird oft auch als echter Diversitätseffekt bezeichnet, weil hier die Leistung von schwachen Genotypen durch die Anwesenheit anderer Genotypen verbessert wird. Das Versuchsdesign von Reusch und seinen Kollegen erlaubte es, zwischen Selektionseffekten und Komplementarität zu unterscheiden [4], denn alle Genotypen, die in Mischungen getestet wurden, lagen auch in Monokulturen vor. Da am Ende des Experimentes auch die gesamten Versuchsflächen erneut genotypisiert wurden, konnte für jeden Genotyp die relative Leistung in Monokultur und Mischung berechnet werden. Die Auswertung zeigte, dass die positive Wirkung der Diversität ausschließlich auf Komplementarität zurückzuführen war, während Selektionseffekte sogar negativ für die gesamten Netto-Biodiversitätseffekte zu Buche schlugen. Negative Selektionseffekte bedeuten, dass die Genotypen, die in Monokultur besonders leistungsfähig sind, in den Mischungen eine unterdurchschnittliche Entwicklung zeigten und sich deutlich langsamer von der Hitzewelle erholten.
Vereinheitlichung ökologischer Theorie
Die neuen Erkenntnisse vereinheitlichen ökologische Theorien. Die Experimente zeigen, dass genotypische Diversität eine ähnliche Funktion übernehmen kann wie Artendiversität in Lebensräumen, die mehrere strukturbestimmende Arten haben. Damit löst sich das scheinbare Paradox von den stabilen und produktiven Monokulturen von Seegräsern, Schilf und Salzmarschen auf. Ob sich Genotypen innerhalb von monodominanten Arten genauso deutlich unterscheiden wie Arten in artenreicheren Lebensgemeinschaften ist eine sich logisch anschließende Frage, die gerade experimentell bearbeitet wird. Die Erkenntnisse zur positiven Wirkung genetischer Diversität sollten auch eine zusätzliche Motivation sein, im angewandten Natur- und Artenschutz nicht allein die Artenvielfalt zu erhalten, sondern gerade auch bei wichtigen Gründerarten wie dem Seegras die genetische Ebene der Biodiversität zu schützen. Darüber hinaus ist zu vermuten, dass genetische Diversität natürlich auch für etwaige evolutive Veränderungen von Populationen als Antwort auf sich ändernde Umweltbedingungen eine entscheidende Voraussetzung ist.